Un modèle est une représentation partielle d’un système que l’on étudie. Il ne prend en compte que les processus et les variables considérés comme pertinents et généralisables pour le modélisateur. Les choix pertinents dépendent des objectifs du modélisateur et la généralisation des phénomènes choisis dépend du domaine d’extrapolation visé.
Les modèles SARRA sont une boîte à outils permettant d’assembler différents modules à partir d’une bibliothèque d’objets évolutive. Malgré leur souplesse, ils restent dans le domaine des modèles classiques de cultures, avec une représentation temporelle quotidienne à l’échelle d’une parcelle. Dans ce modèle, la culture colonise l’espace du sol et de l’air, accède aux ressources disponibles et produit de la biomasse.
Les modèles SARRA-H et SARRA-O sont une version plus détaillée du modèle SARRA, qui est un bilan hydrique dynamique simple utilisé pour estimer l’impact d’un scénario climatique sur une culture annuelle. Dans ces modèles, la plante est considérée comme un conduit couplant une demande atmosphérique (puits) avec une réserve hydrique « utile » dans le sol (source) – un conduit à résistance variable, fonction de la contrainte physique (stress). Les modèles SARRA-H/O supposent que la performance d’une culture est une fonction simple des freins hydriques cumulés au cours d’un cycle végétal.
Ces modèles ont montré une robustesse remarquable à travers les échelles spatiales (de la parcelle à la région) et pour de multiples applications, telles que le système d’alerte précoce de sécurité alimentaire de l’Agrhymet pour les pays du CILSS, l’analyse des risques climatiques locaux et l’estimation des besoins en eau. Cependant, leur validité dépend de la présence d’une forte limitation hydrique et/ou d’une forte variabilité spatiale ou temporelle de cette limitation. Autrement dit, ces modèles se basent entièrement sur une réduction des croissances, mais pas sur la simulation d’un rendement potentiel.
Comparés à d’autres modèles tels que CERES, APSIM et STICS, les modèles SARRA-H/O sont des modèles déterministes et relativement simples, mais simulent plus de processus que AQUACROP, qui est une référence proposée par la FAO. SARRA-H est intégré dans un environnement souple, gérant une librairie de formalismes ainsi qu’une base de données. Ce modèle reproduit des processus à l’échelle de la parcelle permettant des analyses à l’échelle régionale. De plus, il peut être utilisé comme support de base avec d’autres thématiques scientifiques par le biais de modules complémentaires ou d’interfaces avec d’autres modèles (météorologiques, socio-économiques, etc.).
Enfin, la structure modulaire de cet environnement permet une gestion externe du moteur de calcul (simulateur) indépendamment de cet environnement. Toutes ces fonctionnalités sont décrites dans un manuel d’utilisation, et l’environnement offre de nombreux outils et interfaces tels que l’importation de données, l’exportation de données, les représentations graphiques, l’analyse de sensibilité et une base de données pour optimiser la gestion des données et des résultats.
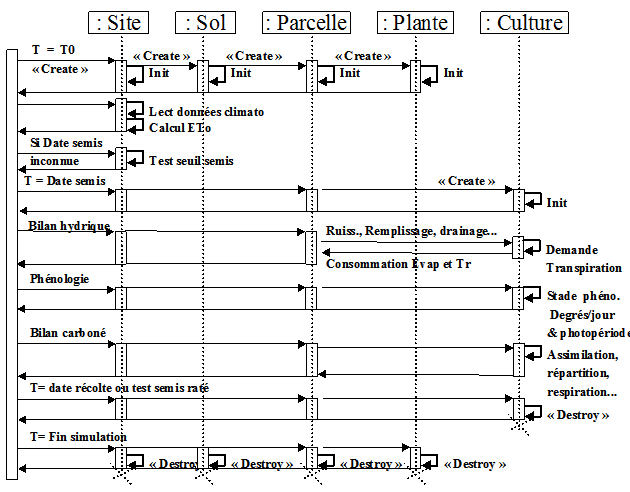
Dans une même boucle journalière le modèle simule trois grands processus
1 : bilan hydrique
il estime l’évapotranspiration et l’indice de stress hydrique; le sol est constitué de trois réservoirs : superficiel, profond et un réservoir racinaire dynamique
2 : bilan carboné
il est basé sur le concept de la grande feuille, il transforme l’énergie solaire en sucre sous contrainte de stress hydrique, puis le répartit dans les biomasses
3: phénologie
lle gère l’évolution des stades phénologiques (émergence, stade végétatif, floraison, maturation) et les processus associés (germination, mortalité juvénile, modalités de répartition des biomasses…)
1 : Bilan hydrique
Concept : une approche de type réservoir
Le bilan hydrique est une approche de type réservoir qui considère le sol comme un ensemble de réservoirs de tailles variables. Chaque réservoir est homogène en termes de processus simulés ou de variables décrivant ses propriétés (infiltration, ruissellement, capacité de stockage, consommation d’eau, etc.). Les flux sont simulés par des processus de remplissage des réservoirs jusqu’à leur capacité maximale (habituellement la capacité au champ), au-delà de laquelle l’eau déborde sur le réservoir suivant. L’eau qui déborde du dernier réservoir est considérée comme du drainage.
Trois types de réservoirs sont retenus pour estimer l’eau disponible pour l’évaporation, la transpiration et le stockage d’eau :
- un réservoir de surface de taille fixe pour gérer l’évaporation du sol ;
- un réservoir en profondeur, sur lequel est simulé l’évolution du front d’humectation pour borner la profondeur maximale des racines, stocker l’eau non accessible par les racines et bloquer l’enracinement dans les sols indurés. Le suivi de l’ensemble de l’eau de la surface à la profondeur maximale est défini dans le stock total.
- un réservoir contenant les racines, dont la profondeur évolue en fonction de la vitesse d’enracinement des racines durant leur développement, pour simuler l’offre en eau pour le processus de transpiration de la plante. Les racines prospectent également le réservoir de surface. Notez que le processus de remontée capillaire n’est pas simulé.
L’équation globale du bilan hydrique est : Stockj+1 = Stockj + apport eau – ruissellement – drainage – (Tr+Evap) Avec : **Tr** : Transpiration réelle (plante zone racinaire), **Evap** : évaporation réelle (demande atmosphérique, réservoir de surface seul), **drainage** (débordement du réservoir profond), **ruissellement** (fn de l'apport d'eau), **eau disponible (infiltrée)** (pluie+irrig-ruissellement), **stock** (stock d'eau dans les réservoirs) .
Apport d’eau & Ruissellement
L’eau disponible (EauDispo) est la somme de la pluie et de l’irrigation nette (IrrigTotDay) moins le ruissellement (Lr). Le ruissellement se déclenche lorsque la pluie dépasse une valeur seuil en mm d’eau (SeuilRuiss), et la quantité d’eau ruisselée dépend d’un pourcentage (PourcRuiss) de la quantité d’eau supérieure au seuil.
Evolution du réservoir racinaire
La taille du réservoir prospecté par les racines évolue en fonction de la vitesse racinaire journalière, définie pour chaque phase phénologique (VRacLevee, VracBVP, VracPSP, VracRPR, VracMatu1, VracMatu2). De plus, deux processus sont pris en compte : l’enracinement en profondeur peut être bloqué par le front d’humectation, et la vitesse d’enracinement journalière peut être réduite en cas de stress hydrique fort (cstr).
Remplissage des réservoirs
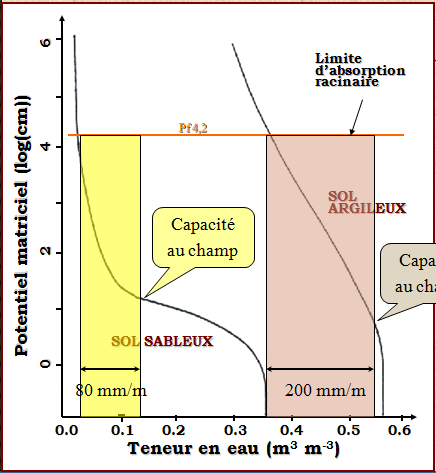
La dynamique de l’eau est représentée par un remplissage par le haut des réservoirs, jusqu’à leur capacité de stockage définie par la capacité au champ (HumCR ou HumFC), et la quantité excédentaire déborde sur le réservoir suivant. La profondeur maximale de l’ensemble des réservoirs est définie, et l’eau qui déborde du dernier réservoir est considérée comme du drainage (Dr). A l’intérieur de chaque réservoir, l’eau est répartie de manière homogène. Les paramètres clés du sol pour ces processus sont la réserve d’eau utile pour la plante (RU), qui est déduite des deux paramètres HumCR et HumPF, représentant respectivement les quantités d’eau à capacité au champ et au point de flétrissement de la plante (Pf 4.2).
Consommation en eau
Le concept d’évapotranspiration est divisé en deux processus, l’évaporation et la transpiration, qui sont étroitement liés au sol et à la plante. Les bases du concept sont similaires à celles de la FAO et utilisent des coefficients de capacité d’évaporation du sol et de transpiration de la plante (Kce et Kcp). Ces processus sont influencés par la relation entre l’offre et la demande d’eau (source/puit) et varient en fonction de la demande atmosphérique (Eto), du taux de couverture et du stade phénologique de la plante. L’évaporation potentielle (evapPot) est limitée par l’eau disponible dans une partie du sol en contact avec l’atmosphère, appelée réservoir de surface, tandis que la transpiration potentielle (TrPot) est limitée par l’eau disponible dans la partie du sol contenant des racines, appelée réservoir racinaire (qui inclut également le réservoir de surface). L’indice de satisfaction hydrique (cstr), qui mesure le rapport entre la transpiration réelle et la demande potentielle de la plante, est utilisé comme frein dans les calculs des biomasses.
- Evaporation (Evap & EvapPot) : elle n’est calculée que sur le réservoir de surface (ex : sol de type sableux avec une capacité de stockage (RU) d’environ 100 mm/m et une profondeur de 20 cm). La profondeur de sol sensible aux relations sol/atmosphère dépend du type de sol (argileux, sableux) une correspondance pourrait se faire en considérant que la variation de profondeur de sol soumis aux effets atmosphériques correspond à une profondeur ayant une capacité de stockage d’environ 200 mm d’eau. Quand le sol est totalement couvert par la végétation on considère qu’il n’y a plus d’évaporation car il y a alors discontinuité entre le sol et l’atmosphère. On traduit ici ce taux de recouvrement (Kce) par le taux de transmission du rayonnement au sol (Ltr : light transmission). Le Ltr est calculé à partir du LAI (Leaf area index, voir Bilan Carbonné). La demande d’évaporation (EvapPot) est égale à ETo x Kce. L’évaporation réelle (Evap) est fonction de l’eau disponible pour la partie évaporation (FESW : Fraction Evaporable Soil Water).
- Note : La densité de semis intervient aussi sur l’évaporation au travers du LAI d’où le Ltr est déduit (Cf paragraphe Prise en compte de la densité de semis dans la partie Bilan carboné)
- Effet mulch et couverture végétale : la couverture végétale (paillis) a plusieurs effets sur le bilan hydrique : 1) stock d’eau dans le sol : réduction du ruissellement mais aussi captation de l’eau par humectation des pailles, 2) réduction de l’évaporation par augmentation du taux de recouvrement. Une série de paramètres caractérisant les effets couvrant, d’humectation, taille des tiges… sont définies par les caractéristique des pratiques culturales (voir fichier SARRA-H V42 parameters2020, publication E. Scopel & al., Modelling crop residue mulching effects on water use and production of maize under semi-arid and humid tropical conditions. Agronomie, EDP Sciences, 2004, 24 (6-7), pp.383-395.10.1051/agro:2004029.hal-00886027 ).
- Transpiration (Tr & TrPot) : elle est calculée sur l’ensemble du réservoir racinaire. Comme pour l’évaporation on déduit la demande potentielle (TrPot) de la demande de l’atmosphère (ETo) et d’un indice de consommation (Kcp) fonction de l’inverse du taux de recouvrement (1-Ltr) et d’un coefficient maximum de transpiration de la plante (KcMax, fn des espèces et qui peut être supérieur à 1). Le stress hydrique de la plante est fonction du PFactor qui permet de définir un seuil de FTSW (Fraction Transpirable Soil Water) simulant le processus d’ouverture ou fermeture des stomates (ex : Pfactor = 0, dès que FTSW est inférieur à 1 fermeture des stomates). Le PFactor est une caractéristique d’espèce et de variétés : par exemple celui du maïs est plus faible que celui du sorgho ou du mil, ce qui rend le mais plus sensible au stress hydrique. La transpiration réelle est calculée en tenant compte du Kcp, de FTSW et du Pfactor.
- Note : La densité de semis intervient aussi sur la transpiration au travers du LAI en effet le Kcp est déduit du Ltr lui même déduit du LAI (Cf paragraphe Prise en compte de la densité de semis dans la partie Bilan carboné)
- Impact du CO2 sur la transpiration (plantes en C4) : la concentration en équivalent CO2 dans l’atmosphère agit sur les processus de régulation des stomates et la demande évaporative de la plante. Cela a été mis en évidence et mesuré sur des cultures de maïs (plante en C4) en plein champ avec apport de CO2 dans l’atmosphère (projet FACE). Cela peut avoir des effets importants en cas de stress hydrique par régulation des stocks d’eau dans le sol : effet de résilience de l’eau dans le sol par réduction de la transpiration des plantes. Ce processus est simulé dans le modèle au travers de deux paramètres : (CO2) qui défini l’équivalent de concentration de CO2 atmosphérique en ppm au niveau du site et SensCO2 qui défini la sensibilité de la plante à cette concentration sur le processus de transpiration.
- Bilan d’eau dans le sol : l’évaporation ne s’effectue que sur le réservoir de surface, alors que la transpiration s’effectue sur le réservoir des racines qui pour partie, ou complètement, intègre le réservoir de surface. On considère que l’évaporation est le processus le plus rapide, on le retranche en premier sur le réservoir de surface. Les estimations d’évaporation et de transpirations ayant été effectuées séparément, on peut avoir une consommation légèrement supérieure à l’eau disponible. On diminuera donc la transpiration en conséquence sous l’hypothèse que le processus d’évaporation est plus rapide.
2 : bilan carboné
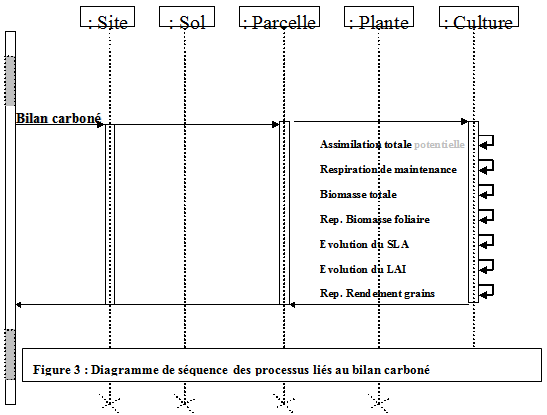
Concept : une approche de type grande feuille

La part photosynthétiquement active, définie par (kPAR), de l’énergie lumineuse interceptée est transformée en assimilats en fonction de l’activité photosynthétique. Cette activité photosynthétique est définie en fonction de la surface foliaire communément traduite par le LAI (« leaf area index ») qui est ici considérée à l’échelle du peuplement d’une parcelle : approche grande feuille (Big Leaf). La fraction de rayonnement interceptée (EpsilonA) est déduite du LAI (loi de Beer). Cette quantité d’énergie permet la création d’assimilas (dBM), en fonction d’un taux de conversion (EpsilonB), ici (TxConversion), est d’une part constant tout au long du cycle et est supérieur à celui communément donné car on simule l’ensemble des assimilats produits alors que le taux de conversion traditionnellement calculé ne tient compte que de l’augmentation de biomasse aérienne sans la biomasse des racines ni la respiration de maintenance (Rm). L’activité photosynthétique peut être contrainte par l’état hydrique et minéral de la plante. Dans le cas d’une contrainte hydrique, le processus de régulation stomatique de la plante intervient.
La répartition des assimilas entre ces biomasses suit des règles d’allocations variant en fonction des phases. Une partie des assimilas est consommée, sans production de biomasse, pour la maintenance des tissus vivants que l’on appelle la respiration de maintenance (Rm). De l’émergence à la floraison, cette biomasse est dissociée en biomasse des racines (BM(rac)) et biomasse aérienne (BM(aér)), elle même dissociée en biomasse foliaire et tige (BM(feuil)) en utilisant des relations allométriques. Après floraison les assimilas disponibles (dBM-Rm) permettent le remplissage des graines.
L’équation globale du bilan carboné est définie par :
BiomasseTotalej+1 = BiomasseTotalej + assimilats - respiration de maintenance. Avec BiomasseTotale = BiomasseRacine + BiomasseFeuilles + BiomassesTiges + BomassesGrains
Evolution du LAI
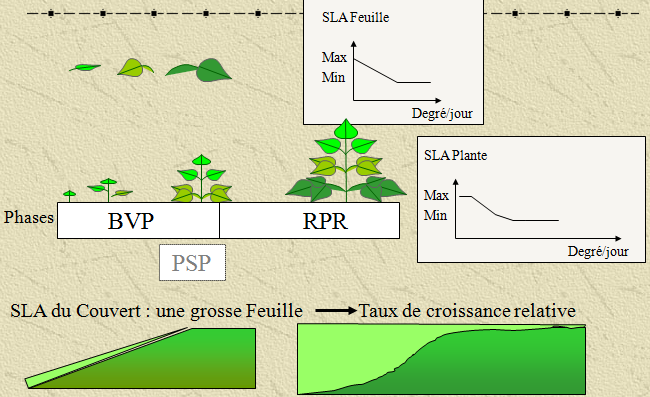
Le LAI (« leaf area index ») est un indice de surface foliaire du peuplement. Il se mesure par le rapport entre la surface des limbes (m²) sur la surface de sol. Le LAI peut donc se déduire de la biomasse foliaire en fonction de la surface massique des feuilles (SLA, specific leaf area). Le SLA décroît en fonction de l’âge de la feuille, dû à son épaississement. D’autres phénomènes interviennent pour l’épaississement de la feuille notamment les niveaux d’exposition au rayonnement et aussi les phénomènes de compétition pour les assimilats. De fait cet indice de surface massique des feuilles est un paramètre clé dans les méthodes de calcul des processus d’accroissement de biomasse s’appuyant sur le LAI pour l’évaluation du processus d’assimilation. D’autre part, sa dynamique varie en fonction des types de plantes (mono et dicotylédones). La surface massique des feuilles est néanmoins un caractère génétique qui peut être définie par des valeurs minimum et maximum. Différents concepts peuvent s’appliquer sur sa dynamique.

Evolution du SLA
les jeunes feuilles ont un SLA supérieur aux vieilles feuilles. La fraction de jeunes feuilles élève le SLA global du couvert. Le paramètre SlaPente provoque une chute générale du SLA de la biomasse foliaire existante (SlaPente = chute relative par jour = fraction de différence entre SLAmax et SLAmin). Ce type de fonctionnement est surtout conçu pour les légumineuses, mais peut être aussi adapté aux autres espèces. Chaque augmentation de la biomasse foliaire journalière est considérée comme des nouvelles feuilles dont la surface massique est égale à SLA(max). La valeur de SLA pour l’ensemble de la biomasse foliaire est alors déduite des moyennes pondérées pour les nouvelles feuilles et les anciennes feuilles (taux de croissance relative).
Rayonnement intercepté (Ltr) : le LAI permet d’évaluer le taux de transmission de lumière au sol par la loi de Beer. On y ajoute un coefficient de « forme » (kdf) qui varie de 0.4 à 0.8 selon la hauteur des plants et l’angle des feuilles (dressées ou non).
Ce coefficient traduit mal les couverts non homogènes.
Assimilation totale potentielle (AssimPot) : La production potentielle d’assimilats est calculée à l’échelle du peuplement sans prise en compte de contraintes hydriques ou minérales. Une relation empirique est définie entre l’activité photosynthétique et le taux de couverture foliaire (1-ltr). On définie un coefficient de conversion de l’énergie lumineuse en matière sèche (KepsiB : kg/ha/j/Mj) constant tout au long du cycle (Txconversion en g/MJ). Seule la part photosynthétiquement active du rayonnement (PAR) est retenue, définie ici par un coefficient kPar (0.5). Un frein hydrique (cstr) est défini par le rapport Tr/TrPot qui réduira l’assimilation potentielle.
Respiration de maintenance journalière (DRespMaint) : la respiration de maintenance varie en fonction de la température et de la biomasse
loi du Q10=2, la valeur double quand la T° augmente de 10°.
La biomasse verte, principalement les feuilles, respire de façon proportionnelle à leur biomasse. Ceci permet d’expliquer le fait que la biomasse plafonne, parfois, même avant la phase de sénescence. Pour calculer les pertes en termes de matière sèche, en supposant qu’il s’agit surtout de CH20, il faut multiplier les valeurs mesurées par 0.65 (rapport poids moléculaire CH20/CO2). Pour la plante entière, il faut prendre en compte les fractions massiques des organes. En supposant 0.03 pour les feuilles et moins de 0.01 pour les autres tissus, on peut faire une estimation de 0.01 pour la plante entière à floraison. La prise en compte du poids plus important de la biomasse foliaire est pris en compte en doublant la valeur définie de TxRespMaint pour cette biomasse.
Remarque : c’est de l’épiaison à la maturité que le poids de la respiration de maintenance est le plus important.
Exemple de valeurs (en g CO2 par g de MS par j):
- mil (feuilles): 0.03 à 25°C
- maïs tempéré (feuilles): 0.032 à 24/18°C
- maïs: 0.026 à 25°C
- riz: 0.02 à 25°C
- sorgho: toute une gamme de résultats entre 0.01 et 0.073, moyenne 0.029 à 30°C
- blé: 0.016 à 20°C
- coton : 0.038 à 30°C
Pour des tissus « non-feuille » on observe des valeurs entre 0.005 et 0.01 (à 20°C pour les tempérés, à 30°C pour les tropicaux).
Augmentation de la biomasse totale (BiomasseTotale & DbiomTot) : la production d’assimilas potentielle est réduite par l’indice de satisfaction en eau de la plante (ie : transpiration réelle/transpiration potentielle). La quantité d’assimilas participant à l’augmentation de biomasse correspond à la quantité nette d’assimilats moins la respiration de maintenance.
Répartition des biomasses : la biomasse totale (BiomasseTotale) est répartie en biomasse racinaire, tige, feuilles et grains
Répartition biomasse totale en racines (BiomasseRacinaire) et biomasse aérienne (BiomasseAerienne) : la répartition en biomasse racinaire et aérienne suit une relation allométrique avant la phase de maturation (KpenteAero, KbaseAero).
Répartition biomasse aérienne en feuilles, tiges puis grains : l’évolution de la biomasse aérienne en biomasse foliaire, de tige et enfin en grains suit deux dynamiques en fonction des phases phénologiques . Avant floraison, on utilise une relation allométrique (cf graphique) qui est une relation stable en cas de stress hydrique. Et après floraison, on se base sur le concept de compétition (répartition prioritaire des assimilas vers les grains). Une sénescence des feuilles peut être favorisée quand l’apport en assimilats (DBiomTot- DRespMaint) est inférieur à la demande. On retranche alors cette différence à la biomasse foliaire et des tiges. La réallocation des réserves est toujours inférieure à la demande en assimilas (fraction KRealloc), ce qui peut s’expliquer par l’énergie dépensée par les processus de réallocation mis en œuvre. Lors du remplissage des grains (capsules, fruits…), la demande journalière de remplissage (DRdtPot) s’ajoute à la respiration de maintenance pour le calcul de la réallocation ce qui se traduit souvent par une scénescence forcée des feuilles pour les variétés ayant un fort potentiel de rendement
- Dans le cas des céréales à partir du stade de l’anthèse (floraison), on considère qu’il n’y a plus d’augmentation de la biomasse végétative (BiomaseVegetative= BiomasseTiges+BiomasseFeuilles).
- Dans le cas des plantes à floraison continue cette relation se poursuit en compétition avec celle liée au remplissage des grains (capsules, fruits…) : cas du coton et du soja. Pour la prise en compte de cette compétition il suffit de spécifier la valeur 6 au paramètre de la variété PhaseDevVeg.

La demande de remplissage des grains : chaque plante a un potentiel de rendement déduit d’une fraction fixe de la biomasse aérienne (potentiel génétique), souvent exprimée par l’indice de récolte (Harvest Index, ex : 30%…50%). En fonction de l’état de la plante, ce potentiel peut être réduit par plusieurs processus : réduction de sites reproducteurs (épis) et du nombre de fruits ou grains. Ces processus ont lieu lors de la phase reproductive, dite sensible (anthèse à floraison). On simule l’effet des contraintes subies par la plante en considérant qu’elles se traduisent par une dynamique moindre d’augmentation de la biomasse totale. Ainsi les contraintes hydriques, induisant une réduction de biomasse lors de cette phase critique, vont se traduire par une réduction du potentiel de rendement, certains le traduisent par le nombre de grains. Ici la réduction du potentiel de rendement est fonction de la réduction de biomasse. Le nouveau rendement potentiel au stade de floraison (RdtPot) est ainsi obtenu par la différence des biomasses aux stades d’initiation paniculaire et de la floraison qui nous donne un indice de pente, avec en base (KRdtPotB) et un coefficient (KRdtPotA) traduisant le potentiel de rendement variétal souvent traduit par l’indice de récolte (Harvest index).
note : on peut retrouver très simplement l’équivalent du nombre de grains juste après floraison en faisant le rapport du (RdtPot) à floraison sur le poid moyen d’un grain (PoidsSecGrain) qui fait partie des variables à définir pour chaque variétés.
Le rendement grain (Rdt) : partant d’un potentiel de rendement (RdtPot), celui-ci dépendra surtout de la phase de remplissage des grains. La contrainte la plus grande est traduite par la satisfaction en eau de la plante où, à ce stade, la demande des grains est la plus forte et considérée comme prioritaire. La demande en biomasse journalière des grains (DRdtPot) est basée sur le concept d’une évolution phénologique en degrés-jours et d’une relation linéaire pour sa dynamique de remplissage des grains, la demande est égale au rapport de la température du jour sur la somme de température définie pour cette phase (Matu1) multipliée par le rendement potentiel.
Pour les espèces à floraison continue (Coton, Soja…), comme à chaque jour on calcul une augmentation du rendement cela s’applique très bien au processus de floraison continue qui peut être bloquée ou s’annulée (phénomène d’abscission pour le coton).
Prise en compte de la densité de semis : en fonction des stratégies de production, des contraintes et des espèces/variétés une densité de semis (en plants/ha) est recommandée (DensOPti).
Nous faisons la double hypothèse : 1) en deçà de cette densité de semis, il n’y a qu’un faible effets de compétitions entre plants une relation de type linéaire est appliquée, 2) au-delà de cette densité recommandée on considère qu’il y a une compétition entre plants (saturation) qui se traduit par une forme asymptotique. Pour tenir compte de ces effets densités sur la dynamique des biomasses et valeurs affectées un simple rapport des densités de semis réelle avec la densité de semis optimale est appliqué à ces valeurs lors des calculs de bilan carbonés.
Ainsi chaque jour, les calculs des processus de bilan carboné (biomasses, rendement et LAI) sont simulés avec une densité optimale. Pour effectuer ces calculs les valeurs de biomasses et LAI sont converti de la densité réelle à cette densité optimale par cette relation asymptotique. Dès que l’ensemble de ces processus sont finalisés, on convertit en sens inverse pour avoir les valeurs rapportées à la densité réelle de semis. Par exemple pour un maïs OPV (open polenisated variety) une densité de semis de 65 000 plants par hectare est conseillée en Afrique ; pour des pratiques intensives avec des variétés hybrides (forte fertilisation, irrigation…) une densité de 85 000 plants/ha est couramment observée en Europe, USA.
Cette relation asymptotique est calibrée au travers de deux autres paramètres : DensiteP pour la pente de la relation, DensiteA pour la borne asymptotique. Cela interagit ainsi sur le LAI, qui est déduit de la biomasse foliaire verte et du SLA rapporté à la plante entière.
- Note : la densité de semis influe donc aussi sur la demande évapotranspirative, dans la partie bilan hydrique, au travers du ltr et du Kcp fonction du LAI qui est fonction de la biomasse foliaire rapportée à la densité de semis réel.
Prise en compte du niveau d’intensification : les objectifs (ou perspectives) de ce modèle sont d’essayer de représenter au mieux la diversité des situations observées en milieu paysan de la parcelle à l’échelle de territoire et notamment pour la petite agriculture familiale. Même dans des conditions assez homogènes le changement d’échelle impact fortement la capacité prédictive des modèles par une absence de connaissance réelle des valeurs et variabilité des facteurs clefs : On a regional scale, the cropping system must be described by only a few variables whose selection depends on the study objective and the involved processes. Collecting cropping system information for all support units is often simplified by identifying determining factors of cropping systems. Obtaining deterministic relations between easily accessible factors and cropping system variables is not always possible, and sometime accessing modalities of determining factors for all support units is also difficult(cf. Leenardt & al., 2010, DOI: 10.1051/agro/2009002)
Dans les milieux tropicaux on est face à une très grande variabilité spatio-temporelle des contraintes et stratégies des agriculteurs. Au sein d’un village on observe des variations de rendements d’un facteur de 10 (de 500 à plus de 5000 kg/ha pour le maïs, au Burkina Faso, voir ci-dessous). Cette variabilité s’observe aussi au sein même des parcelles. Les facteurs intervenants sur cette variabilité sont multiples et en dehors de suivi très fin et fortement contrôlés, il est difficile d’en dissocier et estimer leurs impacts respectif (Projet Sigma et AgMip). Dans un contexte de suivi de parcelles paysannes et plus spécifiquement dans un contexte de petite à toute petite agriculture familiale, il apparait illusoire d’espérer documenter correctement ces différents facteurs à l’échelle des parcelles pour en simuler leurs impacts et encore plus à l’échelle de territoire.
Nous faisons l’hypothèse de traduire l’ensemble des facteurs clefs au sein d’un indice unique dont l’impact sur les processus se traduit au travers du taux de conversion de la variété cultivée à l’exemple de l’effet de la fertilité sur le taux de conversion, **ainsi dans une expérimentation où seul le niveau de fertilité varie il suffit de définir le rapport entre celui qui est observé avec le niveau de fertilité recommandé **.
Dans ce contexte ce rapport est défini par la variable CoeffNI (sup à 0 à plus de 1), qui se doit d’exprimer la combinaison de différents facteurs clefs dont principalement : le niveau de fertilité globale (sol et apports), la qualité du travail du sol dont le contrôle des mauvaises herbes… La valeur 1 de CoeffNI correspond à une situation optimum se référant aux situations recommandées pour la variété. Cette simple relation empirique ne nous permet que de capter la variabilité des rendements observés sur un territoire, comme au Burkina Faso (6 villages répartis dans la province de Tuy, projet Sigma), sans pour autant être capable de l’expliciter. Dans les exemples ci-dessous nous avons défini 4 niveaux d’intensification, passant d’une situation optimum (1), à bonne (0,7), mauvaise (0,3) et très mauvaise (0,1), nous permettant de capter cette variabilité des rendements provenant d’enquêtes à dire de paysans (Mil, Sénégal) ou de mesures sur des parcelles en milieu paysan (Maïs et Sorgho, Burkina). De ces suivis faisant appel à une information très globale on peut ainsi pondérer les résultats obtenus en regard de leur répartition/occurence pour en déduire un rendement réel paysan rapporté à l’échelle d’un territoire.
fig manquante
Pour reproduire ces résultats au travers d’une seule variable définissant ce niveau d’intensification a été établie une double relation asymptotique et une gaussienne inversé permettant d’en déduire le taux de conversion équivalent : TxConversion = NIYo + NIp(1-exp(-NIp NI ))- (exp(-0,5((NI-LGauss)/AGauss)²))/(AGauss2,506628274631). Avec NIYo le décalage en Y de l’asymptote, NIp la pente de l’asymptote, LGauss la largeur de la Gaussienne et AGauss l’amplitude de la Gaussienne. Les quatre niveaux d’intensification définis plus haut, qui ont permis de vérifier la capacité prédictive du modèle au sein de suivis en milieu paysan, sont spécifiés ici par des carrés. Cela nous a permis de calibrer les variables par espèces (cf fichier NI_TxConv.xls).
Phénologie
Traditionnellement le cycle de culture de la plante est découpé en phases de développement, délimitées par des stades phénologiques. Au sein de ces phases les dynamiques d’évolution et/ou l’impact des contraintes sur le développement et le rendement suivent les mêmes lois.
Le cycle de culture peut se diviser en 4 phases principales :
- Phase végétative juvénile (BVP ou « basic vegetative period »)
- Phase sensible à la photopériode (PSP ou « photoperiod-sensitive period »)
- Phase reproductive (RPR ou « reproductive phase … »)
- Phase de maturation (Matu)
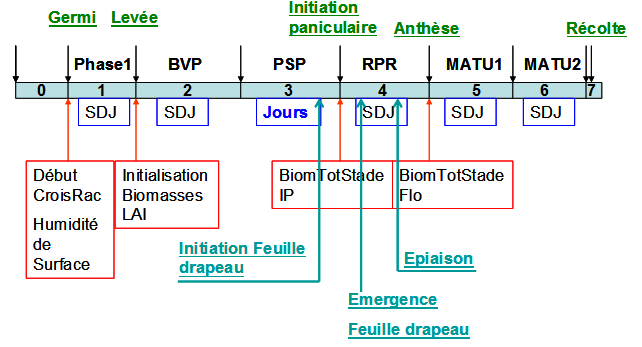
Concept : degrés jours (SDJ) et durée du jour (PP)
Dans SARRA habillée ces phases sont découpées en 7 stades permettant d’optimiser la gestion des méthodes de calcul et de contrôle :
- 0 : avant Semis Durée avant semis (date de semis fixée ou calculée)
- 1 : Germination seuil d’humectation du sol (croissance des racines), puis Durée fonction d’une somme de degrés jour (SDJGermi)
- 2 : BVP Durée fonction d’une somme de degrés jour (SDJBVP) à atteindre
- 3 : PSP Si la variété est photopériodique, durée fonction de la longueur du jour (PP)
- 4 : RPR Durée fonction d’une somme de degrés jour (SDJRPR) à atteindre (nota : phase sensible du stade initiation paniculaire à floraison de l’épiaison à l’anthèse
- 5 : MATU1 Durée fonction d’une somme de degrés jour (SDJMat1) à atteindre, phase sensible de remplissage des grains (Ex : 2/3 phase maturation, grains pâteux)
- 6 : MATU2 Durée fonction d’une somme de degrés jour (SDJMat2) à atteindre, phase non sensible, fin de maturation (dessiccation des grains)
- 7 : Indique le jour de maturation complète permettant la récolte
Lorsque l’on atteint un seuil (somme de température, somme de f(PP)), on change de phase. Ce changement met à jour, en fonction des phases, différentes variables utilisées pour la dynamique de la biomasse. De même en fonction des phases des processus différents seront activés ou désactivés.
Toutes les phases sont définies par une durée en degrés-jour (SumDegreDays), excepté la phase PSP où intervient la photopériode fonction de la latitude :
- Pour les phases fonction d’un cumul de degrés jours le passage à la phase suivante repose sur une somme de température à atteindre (méthode la plus courante et la moins spécifique d’un environnement climatique). Ces seuils de degrés-jours sont déterminés pour chaque variétés.
- La durée de la phase PSP dépend de la sensibilité à la photopériode d’une variété, sa durée varie donc en fonction de la latitude et de la date de semis. La fin de cette phase est toujours marquée par l’initiation paniculaire. La phase RPR est située entre l’initiation paniculaire et se termine à la floraison. La phase maturation se distingue en une sous-phase de remplissage des grains (Matu1) suivie par une sous-phase de dessiccation des grains (grain laiteux à sec). Les phases RPR et une partie de la phase de maturation (Matu1, avant dessiccation) sont les phases les plus sensibles aux contraintes (hydriques…) et ont un impact important sur le rendement dans le cas des céréales.
Les degrés jours : des seuils de températures bornent les étapes de calcul des degrés jour. Tbase température au delà de laquelle la croissance n’est pas bloquée, TL température létale au delà de laquelle la plante ne peut vivre, Topt1 et Topt2 bornes optimum de développement pour la plante. On calcule une vitesse moyenne de développement en considérant qu’elle est linéairement croissante de 0 à 1 entre Tbase et Topt1, puis constante et égale à 1 entre Topt1 et Topt2 puis linéairement décroissante de 1 à 0 entre Topt2 et Tlethale. On calcule alors la température équivalente comprise entre Tbase et Topt1 donnant la même vitesse de développement. Les degrés jours correspondent à cette température équivalente.
La photopériode : une photopériode (PP) défavorable ralentie le processus de l’induction florale. La durée de cette période d’induction, normalement instantanée sous conditions favorables, est fonction d’une valeur seuil les concepts et méthodes de calculs sont décrites dans une publication (M. Dingkhun et al., 2008). L’initiation florale se déclenche quand ce seuil critique est atteint.  La PP utilisée dans le modèle est la durée astronomique du jour, sans tenir compte du « civil twilight ». Dans le « civil twilight », la durée du jour est plus longue car elle intègre les périodes faiblement éclairées avant l’apparition et la disparition du soleil, où la plante est déjà physiologiquement réactive
La PP utilisée dans le modèle est la durée astronomique du jour, sans tenir compte du « civil twilight ». Dans le « civil twilight », la durée du jour est plus longue car elle intègre les périodes faiblement éclairées avant l’apparition et la disparition du soleil, où la plante est déjà physiologiquement réactive